En 1935, von Huene décrivit un squelette partiel (NHMUK PV R747) découvert dans le Rhétien de la formation géologique de Westbury (Somerset, Angleterre). Elle en fit l’holotype du nouveau genre Pachystropheus, avec P. rhaeticus pour espèce. von Huene classa alors Pachystropheus comme un choristodère, un groupe de sauropsides alors considéré comme faisant partie de Rhynchocephalia. Ce genre fut longtemps resté hors de l’attention des paléontologues, sans faire l’objet de révisions. A noter qu’en 1961, Kuhn attribua sa propre famille à Pachystropheus : Pachystropheidae.

En 1996, Storrs et ses collègues redécrivirent Pachystropheus et confirmèrent son classement au sein de Choristodera. Ils notèrent également que le genre Rysosteus, décrit par Owen en 1842 pour un squelette fragmentaire (aujourd’hui perdu) de la formation géologique de Westbury (Gloucestershire, Angleterre), pouvait être synonymisé à Pachystropheus rhaeticus. En 2005, Renesto a suggéré que Pachystropheus puisse être un thalattosaure proche d’Endennasaurus. Quinn et ses collègues redécrivent ainsi Pachystropheus afin de mieux connaître son classement et d’établir une liste des spécimens correctement attribué à ce taxon.

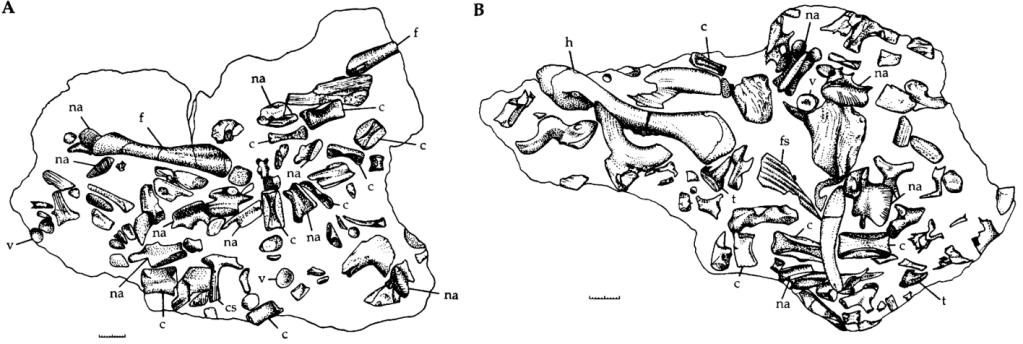
L’holotype (NHMUK PV R747) de Pachystropheus rhaeticus est un assemblage d’éléments postcrâniens désarticulés composé de 35 vertèbres, de côtes, des iliums, du fémur gauche et d’une fibula. Quinn et ses collègues indiquent que cet assemblage représente probablement plus d’un individu. De plus, plusieurs éléments assignés à NHMUK PV R747 par von Huene (1935) ou Storrs et al. (1996) sont en réalité des os de cœlacanthes associés aux os de Pachystropheus.

Il existe de nombreux spécimens historiquement référés à Pachystropheus. Dufin (1978 et 1980), Martill et Down (1986), Storrs et Gower (1993), Storrs et al. (1996), Nordén et al. (2015), Mears et al. (2016), Cueille et al. (2020), Čerňanský et al. (2020), Moreau et al. (2021) et Cawthorne et al. (2024) (voir cet article) ont attribué à P. rhaeticus des spécimens de la formation géologique de Westbury (Angleterre et Pays de Galles). Seulement une partie de ces spécimens ont été révisés par Quinn et ses collègues
Des spécimens d’autres localités d’Europe ont également été assignés à Pachystropheus. Sander et al. (2016) lui ont attribué des spécimens de la formation géologique d’Exter (Allemagne). Bardet et al. (1999) lui ont attribué des spécimens du gisement du Pic des Courmettes (Alpes Maritimes, France). Enfin, Delsatte (2002) lui a référé des spécimens de la formation géologique du Grès de Mortinsart (Belgique et Luxembourg). Quinn et ses collègues n’ont toutefois pas révisé l’attribution de ces spécimens.

Tous ces spécimens représentent des éléments des membres ou du squelette axial, mais aucun spécimen crânien représentant avec certitude Pachystropheus n’a été décrit. Parmi ces spécimens historiquement référés à Pachystropheus, Cueille et al. (2020) ont attribué deux vertèbres caudales (BRSMG Cf15467) à P. rhaeticus. La particularité de BRSMG Cf15467 (mal noté BRSMG Cf15546 dans certaines sections de l’article) est qu’il s’agit d’un coprolithe spiralé, probablement produit par un chondrichtyen. Čerňanský et al. (2020) ont également décrit l’ostéohistologie de P. rhaeticus à partir d’un fémur droit (Z 28225).
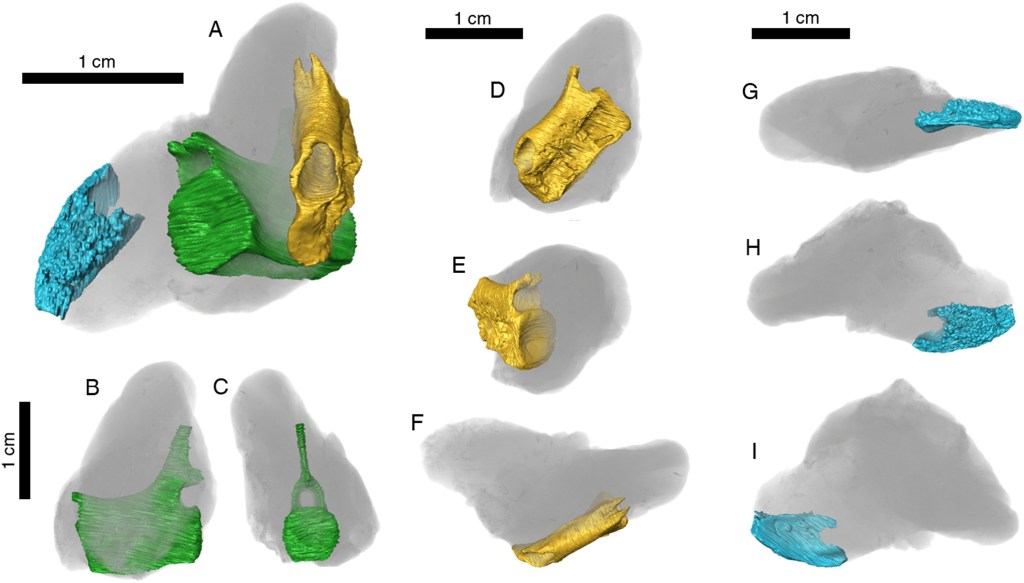
Quinn et ses collègues réfèrent également de nouveaux spécimens à P. rhaeticus. Les plus complets d’entre eux sont un assemblage d’éléments postcrâniens désarticulés (BRSMG Cg3102) composé de 62 vertèbres, des côtes, des humérus, des fémurs, un radius, un tibia, des phalanges et des éléments non identifiés et un assemblage d’éléments postcrâniens désarticulés (BRSMG Ce10791) composé d’éléments des membres, de vertèbres et de côtes.

Quinn et ses collègues réfèrent à P. rhaeticus un humérus associé à un fémur (NMW 2001.42G.563), des éléments des membres associés (BRSMG.Ce17082-17094), un fémur, des côtes et des vertèbres associés (STGCM 2008.50_1089), des côtes associées à un humérus (BRSMG Ce10788), des côtes associées (BRSMG Cd3868), 4 spécimens avec des vertèbres et des côtes associées (BRSMG Ce17860, STGCM 06.62/2, BRSMG Ce17785 et BRSMG Ce17771), un fémur associé à un radius (BRSUG.25304), une phalange associée à une vertèbre caudale (BRSMG Ce17656) ainsi qu’un fémur associé à des vertèbres (BRSMG Ce17770).

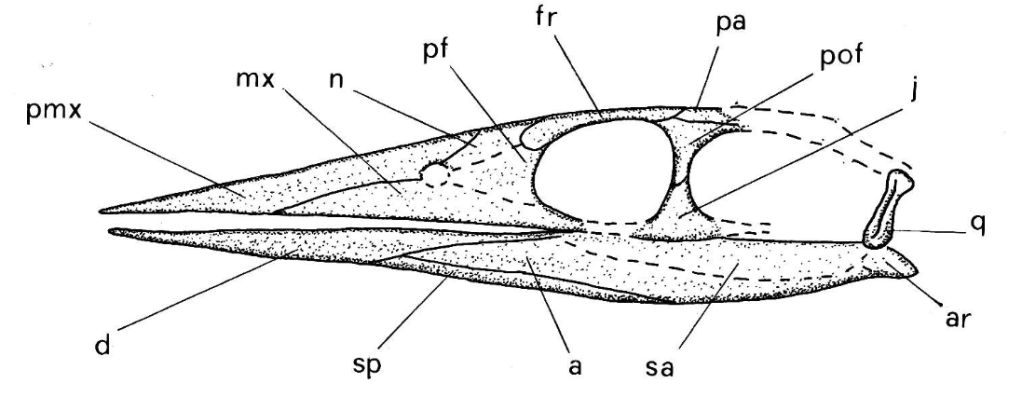
Ils lui réfèrent un jugal (BRSMG Ce17375), le seul élément crânien actuellement connu pour ce taxon. Ils lui attribuent de nombreuses vertèbres isolées (NMW 2001.42G.617, NMW 2001.42G.986, STGCM 1986.145/1-8, STGCM 1986.145, STGCM 1944.115, BRSUG.25312, BRSUG.25302-2, BRSUG.25305, BRSMG Ce17854, BRSMG Ce17789-17790, BRSMG Ce4878-4886, BRSMG Cf650-652, BRSMG Ce17799, BRSMG Ce17142-17182, BRSMG Ce16801, BRSMG Glos.305218, BRSMG Ce17340, BRSMG Ce10789, BRSMG Cb4879, BRSMG Cb8929, BRSMG Ce16914, BRSMG Ce10779 et BRSMG Cd2705), une gastralia (BRSMG Ce17801), deux côtes isolées (BRSMG Ce17859 et NMW 2001.42G.673)

Au niveau des éléments du squelette appendiculaire, Quinn et ses collègues attribuent à P; rhaeticus deux clavicules isolés (BRSMG Ce17080 et NMW 2002.57G.9), une interclavicule (BRSUG.19969), un élément du bassin ou de la ceinture scapulaire (BRSMG Ce16957), deux iliums isolés (BRSUG.25318 et NMW 2015.30G.24), cinq éléments indéterminés de membre (STGCM 1960/62/3, BRSMG Ce17607, NMW 2001.42G.675, BRSMG Ce17794 et BRSMG Ac82-71), deux phalanges isolés (BRSMG Cd2064 et NMW 2001.42G.984), un unguéal (BRSMG Ce17199) et un fragment osseux (BRSMG Ce10782).

Les éléments les plus représentés de Pachystropheus sont les humérus et les fémurs. Quinn et ses collègues lui attribuent 22 humérus (NMW 28.2.G231,G232, NMW 2001.42G.73-74, NMW 2001.42G.610, STGCM 1986.52, BRSUG.25341, BRSUG.7006, BRSMG Ce17642, BRSMG Ce17772, BRSMG Ce17793, BRSMG Ce17201, BRSMG Ce17323, BRSMG Ce17191, BRSMG Ce17192, BRSMG Ce16808-16811, BRSMG Ce16862, BRSMG Cd2061, BRSMG Ac82-16, BRSMG Cf3206, BRSMG Cd3365 et BRSMG Cd4050) et 21 fémurs (NMW 2001.42G.78, NMW 2001.42G.80-81, NMW 2001.42G.560, NMW 2001.42G.565, NMW 2001.42G.608, NMW 2001.42G.667, NMW 2001.42G.751, NMW 2001.42G.708, STGCM 1968.2, BRSMG Ce17645, BRSMG.Ce17643-17644, BRSMG Ce17330, BRSMG.Ce17791, BRSMG Ce17336, BRSMG Ce17641, BRSMG Ce17342, BRSMG Ce16920, BRSMG Ce7322, BRSMG Ce16854, BRSMG Ce10798 et BRSMG Cd2059).

L’analyse phylogénétique de Quinn et ses collègues classe Pachystropheus au sein de Thalattosauria, ce qui confirme les hypothèses de Renesto (2005) et Cawthorne et al. (2024). Au sein de Thalattosauria, Pachystropheus est classé en taxon-soeur d’Endennasaurus chez les Askeptosauroidea. Bien que Quinn et ses collègues ne le suggèrent pas, il est possible qu’Endennasaurus et Pachystropheus forment une famille de thalattosaures askeptosauroidés qui pourrait prendre le nom déjà disponible de Pachystropheidae.

Malgré la grande abondance de spécimens de Pachystropheus, la plupart d’entre eux sont désarticulés et manquent cruellement de matériel crânien et de dents. Quinn et ses collègues expliquent ce manque de crânes par la possibilité que le crâne de Pachystropheus soit similaire à celui de son proche parent Endennasaurus. Pachystropheus aurait ainsi eu un crâne gracile aux mâchoires fines, allongées et édentées. Cela en aurait fait un prédateur spécialisé dans la chasse aux invertébrés marins au corps mou comme les céphalopodes par exemple.
Le très grand nombre de spécimens de Pachystropheus découverts dans une zone restreinte peut laisser penser que ce taxon était très abondant, et qu’il vivait potentiellement en colonies. Il s’agissait d’un prédateur d’environ 1 mètre de longueur mais pouvant parfois atteindre jusqu’à 2,5 mètres de longueur. Ses os denses et épais ainsi que son squelette témoignent d’un mode de vie aquatique pour Pachystropheus. Ses membres robustes suggèrent toutefois qu’il était capable de se mouvoir sur terre, contrairement à d’autres thalattosaures.
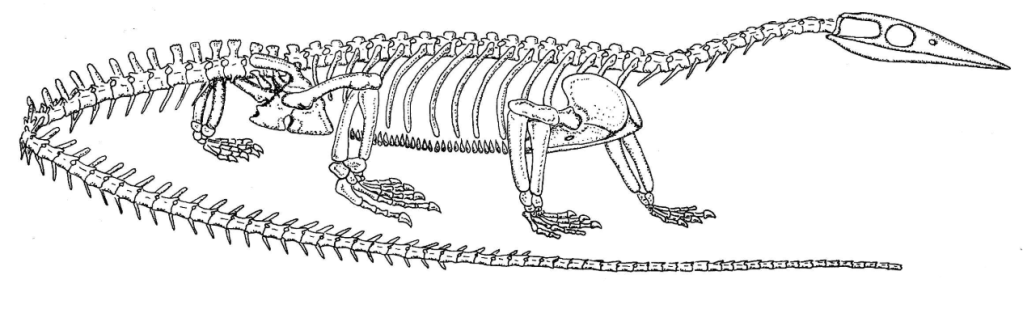
La désarticulation des spécimens de Pachystropheus s’explique par le fait que ces spécimens ont été déposés lors de tempêtes. Pachystropheus vivait dans un environnement côtier subtropical, probablement lagunaire, à proximité d’un archipel d’îles. Il vivait en compagnie de plésiosaures, d’ichthyosauromorphes et de placodontes. C’était un prédateur intermédiaire pouvant être la proie de requins, de gros poissons téléostéens, de cœlacanthes ou d’ichthyosaures.

Références : Quinn, J.G.; Matheau-Raven, E.R.; Whiteside, D.I.; Marshall, J.E.A.; Hutchinson, D.J. ; Benton, M.J., 2024, The relationships and paleoecology of Pachystropheus rhaeticus, an enigmatic latest Triassic marine reptile (Diapsida: Thalattosauria). Journal of Vertebrate Paleontology. e2350408.
von Huene, E., 1935, Ein Rhynchocephale aus dem Rhät. Neues Jahrbuch für Mineralogie, Geologie und Paläontologie, Abteilung B. 74: 441-447.
Kuhn, O., 1961, Fossilium Catalogus, I: Animalia, pt. 98, Reptilia Supplementum I (Millerosauria, Placodontia, Eunotosauria, Bolosauria, Weigeltisauria, Thalattosauria, Pleurosauria, Araeoscelidia, Choristodera). W. Junk, ‘s-Gravenhage.
Storrs, G.W.; Gower, D.J.; Large, N.F., 1996, The diapsid reptile, Pachystropheus rhaeticus, a probable choristodere from the Rhaetian of Europe. Palaeontology. 39: 323-349.
Owen, R., 1842, Report on British fossil reptiles. Part II. Report of the Eleventh Meeting of the British Association for the Advancement of Science. 1842: 60-204.
Renesto, S.C., 2005, A possible find of Endennasaurus (Reptilia Thalattosauria), with a comparison between Endennasaurus and Pachystropheus. Neues Jahrbuch für Geologie und Paläontologie. 2005(2): 118-128.
Cawthorne, M.; Whiteside, D.I.; Benton, M.J., 2024, Latest Triassic terrestrial microvertebrate assemblages from caves on the Mendip palaeoisland, S.W. England, at Emborough, Batscombe and Highcroft Quarries. Proceedings of the Geologists’ Association.
Duffin, C.J., 1978, The Bath Geological Collections. f. The importance of certain vertebrate fossils collected by Charles Moore: an attempt at scientific perspective. Geological Curator. 2(2): 59-67.
Duffin, C.J., 1980, The Upper Triassic section at Chilcompton, Somerset, with notes on the Rhaetic of the Mendips in general. Mercian Geologist. 7: 251-268.
Martill, D.M.; Dawn, A., 1986, Fossil vertebrates from new exposures of the Westbury Formation (Upper Triassic) at Newark, Nottinghamshire. Mercian Geologist. 10: 127-133.
Storrs, G.W.; Gower, D.J., 1993, The earliest possible choristodere (Diapsida) and gaps in the fossil record of semi-aquatic reptiles. Journal of the Geological Society. 150(6): 1103-1107.
Nordén, K.K.; Duffin, C.J.; Benton, M.J., 2015, A marine vertebrate fauna from the Late Triassic of Somerset, and a review of British placodonts. Proceedings of the Geologists’ Association. 126(4-5): 564-581.
Mears, E.M.; Rossi, V.; MacDonald, E.; Coleman, G.; Davies, T.G.; Arias-Riesgo, C.; Hildebrandt, C.; Thiel, H.; Duffin, C.J.; Whiteside, D.I.; Benton, M.J., 2016, The Rhaetian (Late Triassic) vertebrates of Hampstead Farm Quarry, Gloucestershire, UK. Proceedings of the Geologists’ Association. 127(4): 478-505.
Cueille, M.; Green, E.; Duffin, C.J.; Hildebrandt, C.; Benton, M.J., 2020, Fish and crab coprolites from the latest Triassic of the UK: From Buckland to the Mesozoic Marine Revolution. Proceedings of the Geologists’ Association. 131(6): 699-721.
Čerňanský, A.; Klein, N.; Renesto, S.; Michalik, J.; Šurka, J.; Sentpetery, M., 2020, The first skeletal evidence of an Upper Triassic (Rhaetian) diapsid reptile from the Western Carpathians (Strážov Highlands, Slovakia). Geologica Carpathica. 71(2): 134-149.
Moreau, M.; Duffin, C.J.; Hildebrandt, C.; Hutchinson, D.; Parker, A.; Carpenter, S.; Benton, M.J., 2021, Microvertebrates from the Rhaetian basal bone bed of Saltford, near Bath, SW England. Proceedings of the Geologists’ Association. 132(2): 174-187.
Sander, P.M.; Wintrich, T.; Schwermann, A.H.; Kindlimann, R., 2016, Die paläontologische Grabung in der Rhät-Lias-Tongrube der Fa. Lücking bei Warburg-Bonenburg (Kr. Höxter) im Frühjahr 2015. Geologie und Paläontologie in Westfalen. 88: 11-37.
Bardet, N.; Cuny, G.; Lachkar, G.; Pereda Suberbiola, X.; Mazin, J.M.; Thomel, G.; Barbieri, L.; Vasse, D.; Parot, B.; Emily, A., 1999, A new marine vertebrate fauna from the uppermost Triassic of Eastern Provence (France). In: Jagt, J.W.M.; Lambers, P.H.; Mulder, E.W.A.; Schulp, A.S. (eds.), Proceedings of the Third European Workshop on Vertebrate Palaeontology, Maastricht, 6–9 May, 1998. Geologie en Mijn bouw. 78(3–4): 12.
Delsate, D., 2002, Présence du diapside Pachystropheus rhaeticus dans le Trias supérieur en faciès rhétien de Markusbierg-Burmerange (Grand-Duché de Luxembourg) et de Habay-la-Vieille (Belgique). Archives-Institut Grand-Ducal de Luxembourg. Section des Sciences Naturelles, Physiques et Mathématiques. 44: 15-23.
Renesto, S., 1992, The anatomy and relationships of Endennasaurus acutirostris (Reptilia: Neodiapsida) from the Norian (Late Triassic) of Lombardy. Rivista Italiana di Paleontologia e Stratigrafia. 97: 409-430.
Toutes les images proviennent de Quinn et al., 2024, à l’exception de la première et de la quatrième qui proviennent de Storrs et al., 1996, de la deuxième qui provient de la galerie en ligne du Muséum National d’Histoire Naturelle du Royaume Uni, de la sixième qui provient de Cueille et al., 2020, de la huitième qui provient de la galerie en ligne du Muséum de Bristol ainsi que de la treizième et de la quatorzième qui proviennent de Renesto, 1992
