Le caryotype est l’arrangement des chromosomes d’une cellule chez une espèce, dont l’analyse est grandement facilitée par les études cytogénétique et cytogénomiques du génome. Ces études permettent de cartographier le caryotype et de connaître la forme des chromosomes par exemple. Le caryotype des espèces actuelles est connu grâce à son échantillonnage sur de nombreux individus, mais celui d’espèces disparues ne peut être connu car l’ADN ne se préserve pas lors de la fossilisation. Plusieurs études ont déjà été réalisées pour reconstruire le caryotype de groupes fossiles, notamment O’Connor et ses collègues en 2018. Griffin et ses collègues cherchent ainsi à déterminer le caryotype des dinosaures non-aviens en se basant sur cette étude ainsi que sur le caryotype d’espèces d’oiseaux, de lepidosaures, de tortues et de crocodiliens actuels.
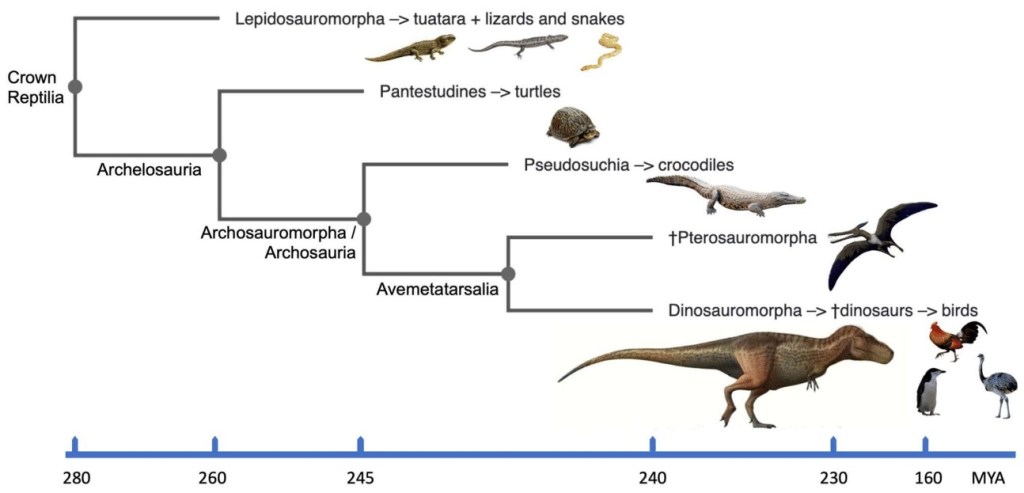
Grâce aux analyses moléculaires et à la phylogénie, il a pu être déterminé que le groupe actuel le plus proche des dinosaures est celui des oiseaux, puisque ceux-ci sont eux même des dinosaures. Ensuite, ce sont les crocodiliens qui leur sont les plus proches, avec un ancêtre commun au sein d’Archosauria, puis les tortues qui partagent un ancêtre commun avec les dinosaures au sein d’Archosauromorpha. Finalement, le groupe le plus éloigné par Griffin et ses collègues, les lepidosaures, partagent un ancêtre commun avec les dinosaures au sein de Sauria. L’époque pendant laquelle ces différents groupes sont apparus et ont divergé des autres est inconnue, mais se situe probablement au début du Permien, vers -280 millions d’années.
D’après Griffin et ses collègues, le caryotype du rhynchocéphale actuel Sphenodon (le tuatara) comporte 36 chromosomes (à noter que tous les nombres de chromosomes donnés dans cet articles valent pour 2n, c’est-à-dire en phase diploïde), avec 14 paires de macrochromosomes et 4 paires de microchromosomes (macrochromosomes et microchromosomes ne se distinguent que par leur taille, les microchromosomes étant bien plus petits). Son caryotype est très similaire à celui de la plupart des tortues actuelles et suggère que le caryotype ancestral des Sauria comportait probablement 14 paires de macrochromosomes et un nombre variable de paires de microchromosomes.

Pour ce qui est du caryotype des squamates, celui-ci est assez variable avec entre 16 et 62 chromosomes. Le caryotype des crocodiliens est constitué de 32 macrochromosomes, mais pas de microchromosomes, ce qui est une exception au sein des Sauria actuels. Les oiseaux possèdent quant à eux une moyenne de 80 chromosomes, avec une majorité de microchromosomes.
En 2018, O’Connor et ses collègues avaient déjà déterminé le caryotype ancestral de plusieurs groupes. Griffin et ses collègues reprennent donc leurs résultats et établissent le caryotype ancestral des Sauria à entre 36 et 46 chromosomes, avec une moitié de microchromosomes et une autre de macrochromosomes, qui remonte donc à -280 MA. Sur la base de l’étude d’O’Connor et ses collègues, Griffin et ses collègues montrent que le caryotype des premiers archosauromorphes comportait 66 chromosomes et que celui-ci n’a ensuite pas subi de réarrangement majeur au cours de l’évolution. Les pseudosuchiens présentent la particularité d’avoir perdu leurs microchromosomes au cours de leur évolution, avec un caryotype ne comportant plus que 34 chromosomes.
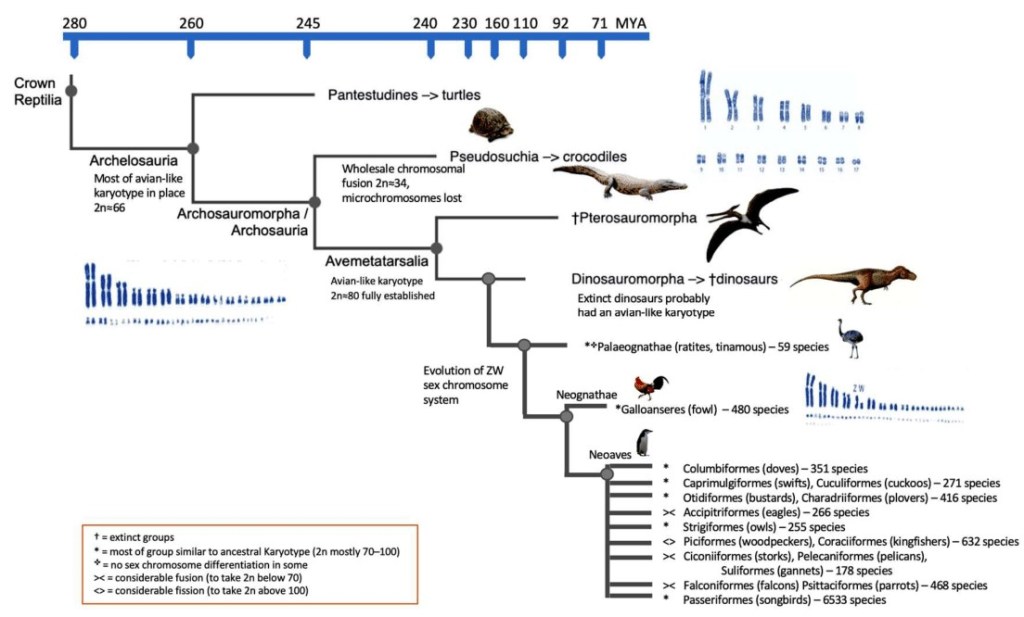
Le caryotype des oiseaux, avec 80 chromosomes en moyenne, a probablement vu le jour après la divergence des pseudosuchiens, c’est-à-dire au sein d’Avemetatarsalia. Cela implique le fait que les ptérosaures et les dinosaures avaient probablement un caryotype similaire. Le système des chromosomes sexuels W et Z que l’on observe chez les oiseaux néognathes ne se retrouve pas chez les oiseaux paléognathes ni chez les autres membres de Sauria. Selon Griffin et ses collègues, il est improbable que les dinosaures non-aviens aient eu des chromosomes sexuels, bien qu’il ne soit pas impossible que ce trait soit apparu chez les avemetatarsaliens et que ce trait ait ensuite disparu chez les paléognathes.

Griffin et ses collègues ont noté le manque de différences majeures entre le caryotype des tortues et celui des oiseaux. Cela indique que le modèle chromosomique des oiseaux est resté pratiquement inchangé depuis des centaines de millions d’années et était presque identique chez les dinosaures non-aviens et les ptérosaures. Ce manque de changement de modèle contraste avec la grande diversité phénotypique des dinosaures. Cette diversité peut s’expliquer par la présence de nombreux chromosomes et par de nombreuses altérations interchromosomiques. Par ailleurs, le fait que le modèle chromosomique des oiseaux est probablement antérieur à l’apparition des dinosaures s’ajoute aux plumes à la pneumatisation du squelette qui sont des caractères typiques des oiseaux dont l’origine remonte à avant l’apparition des dinosaures.
Références : Griffin, D.K.; Larkin, D.M.; O’Connor, R.E.; Romanov, M.N., 2022, Dinosaurs: Comparative Cytogenomics of their Reptile Cousins and Avian Descendants. Animals. 13: 106.
O’Connor, R.E.; Romanov, M.N.; Kiazim, L.G.; Barrett, P.M.; Farré, M.; Damas, J.; Ferguson‐Smith, M.; Valenzuela, N.; Larkin, D.M.; Griffin, D.K., 2018, Reconstruction of the diapsid ancestral genome permits chromosome evolution tracing in avian and non‐ avian dinosaurs. Nature Communications. 9: 1883.
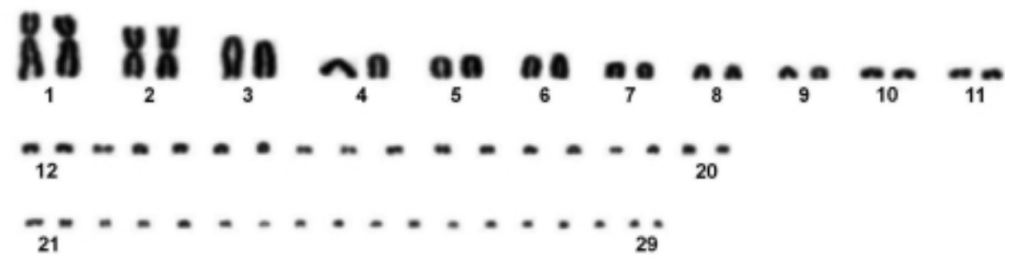
Noleto, R.B.; Kantek, D.L.Z.; Swarça, A.C.;Dias, A.L.; Fenocchio, A.S.; Cestari, M.M., 2006, Karyotypic characterization of Hydromedusa tectifera (Testudines, Pleurodira) from the upper Iguaçu River in the Brazilian state of Paraná. Genetics and Molecular Biology. 29(2): 263-266.
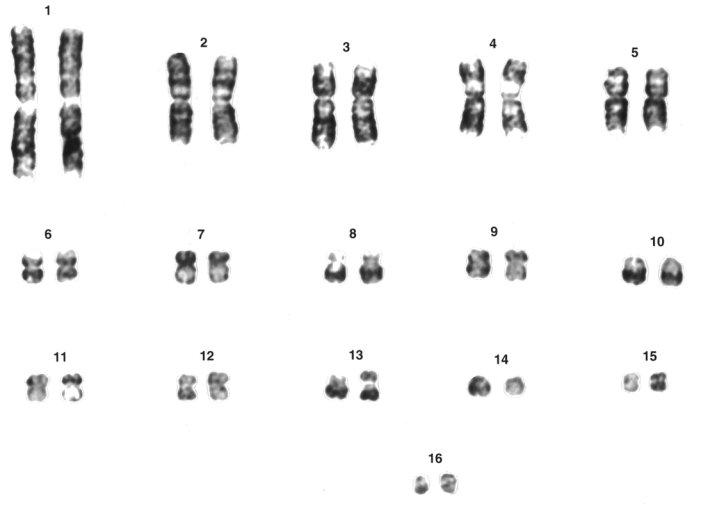
Castiglia, R.; Flores-Villela, O.; Bezerra, A.; Muñoz A.; Luis, A.; Gornung, E., 2013, Pattern of chromosomal changes in ‘beta’ Anolis (Norops group) (Squamata: Polychrotidae) depicted by an ancestral state analysis. Zoological Studies. 52: 60.
Valleley, E.M.A.; Harrison, C.J.; Cook, Y.; Ferguson, M.W.J.; Sharpe, P.T., 1994, The karyotype of Alligator mississippiensis, and chromosomal mapping of the ZFY/X homologue, Zfc. Chromosoma. 103: 502-507.
Ji, M.; Guan, W.; Gao, Y.; Li, L.; Bai, C.; Ma, Y.; Li, X., 2016, Cultivation and Biological Characterization of Chicken Primordial Germ Cells. Brazilian Archives of Biology and Technology. 59: e16150374.
Toutes les images proviennent de Griffin et al., 2022 à l’exception de la première qui provient de Noleto et al., 2006, de la troisième qui provient de Castiglia et al., 2013, de la quatrième qui provient de Valleley et al., 1994 ainsi que de la dernière qui provient de Ji et al., 2016
